webbtidskrift för historia och kultur
Du är här: >> Avdelning Historia >>Paleoantropologi
Den första människan med talets gåva
En arkeosofisk essä på tvärvetenskaplig grund om Homo heidelbergensis
av Sören G Lindgren
Kapitel 7
Klimatchockerna
1. Skärgårdens Europa | 2. Tiden i Afrika | 3. I sandtagen | 4. ”Fadern” och syskonen | 5. I Afrika och Levanten | 6. I Europa | 7. Klimatchockerna | 8. Första språket | 9. Denisovernas konst | 10. Strecket som symbol | 11. Sociala utvecklingen | 12. Språktraditionen | 13. Kulturarvet
ad var det som gjorde att människan blev ”skapelsens krona” som man sade förr i en projicering av feodalismens världsbild på människans evolutionära utveckling. Svaret av i dag lyder helt enkelt: Det var miljön. Där omgivningarna var stabila genom miljoner år som i Sydostasien, där gibbonerna obetydligt utvecklats, eller i centrala Afrika, där gorillor och schimpanser inte heller uppvisar några stora evolutionssteg, där skedde ingen evolution av betydelse.

Charles Darwin gjorde en viktig resa runt jorden 1831-36, varvid han fick idén till det naturliga urvalet. Efter mycket funderande publicerade sin banbrytande bok ”Om arternas uppkomst” år 1859 (senaste svenska översättning år 1980). Bild från Wikipedia.
Men runt om det tropiskt skogiga Afrika genomgick miljön täta och dramatiska förändringar. Av de djurarter som klarade påfrestningarna skulle en bli Homo sapiens.
Av de tidiga undersökarna av evolutionen var det Alfred Russel Wallaces som mest betonade miljöns roll. Wallace (1823-1913) var verksam samtidigt med Charles Darwin (1809-1882).
Sedan slutet på 1700-talet hade den tanken börjat slå rot att inte bara bergen och över huvud landskapen utan även allt levande haft en lång utveckling bakom sig. Men ingen kom på hur detta kunde ha skett.

Alfred Russel Wallace fotograferad på gamla dagar då han slagit sig ned i London för gott. Han försökte aldrig konkurrera med Darwin utan ställde sig alltid stöttande till och solidarisk med denne. Först under senare år har han fått erkännande som självständig tänkare. Foto London Stereoscopic & Photographic Company (aktivt 1855-1922).
Wallace tänkte på egen hand fram en förklaring under sin verksamhet i den indonesiska övärlden. Han menade att ett naturligt urval förde utvecklingen framåt men ansåg också att det var miljön, som inte bara drev på utan även kontrollerade och styrde evolutionen (Wallace 1858).
Darwin var inte främmande för tanken på miljöns inflytande men viktigare var hans förslag om konkurrens mellan individer av en art om föda och sexualpartners, ty det förklarade processen mera i detalj genom att tilldela individerna ett eget spelrum (Darwin 1859).
De flesta paleoantropologer betygar sin vördnad för Darwin i något sammanhang. Men i praktiken är det Wallace tankar om miljöinflytandets avgörande roll som kommit att ta överhanden under senare år. Till denna utveckling bidrog den framstående finländska paleontologen Björn Kurtén (1924-88), som öppnade för ett ekologiskt tänkande inom paleontologin på 1960- och -70-talen.
Björn Kurtén

Björn Kurtén var professor i paleontologi vid Helsingfors universitet. Som docent motarbetades han av äldre kolleger, som anåg att hans matematiska beräkningar inte tillhörde deras vetenskap. När han började få internationellt erkännande tilldelades han en personlig professur år 1972.
Ty numera vet vi att generna på en mängd olika sätt håller sig underrättade om vad som händer i miljön. Viktigast är kanske de enzym som kallas protein kinases (Lai et al 2016) och som bl a förmedlar information mellan cellens olika delar men också uppgifter till och från andra celler och därmed även från miljön. Även om cellen inte har ett medvetande som vårt, som kan göra bedömningar av vädret en given dag och därmed få oss att välja klädsel, så svarar generna på den inkomna informationen, men troligen på ett ganska stelt, mekaniskt sätt. Det gör att genomet får problem när miljön snabbt förändras (i fallet med Lucys ättlingar ledde det till utdöd, detsamma drabbade Paranthroperna (en till ett liv på slätterna anpassad australopitecin)). Ännu värre blir det när klimatomslagen kommer tätt.
Vad som bidrog till att framkalla klimatomslagen var istiderna eller glaciationerna. Men det kan vara mycket jobbigt att komma ihåg namnen de många glaciationerna för mycket länge sedan. Vad som är viktigt är att veta att Antarktis var täckt av inlandsis från för 2,6 miljoner år sedan och att istiderna i början var 40 000 år långa för att för omkring 1 miljon år tillbaka oförklarligt bli mer än det dubbla (cirka 100 000 år). Sålunda började den senaste s k Weichsel-glaciationen för 115 000 år tillbaka (och slutade för omkring 11 000 år sedan). Men för den läsare som är villig att fördjupa sig i de mycket gamla glaciationerna har jag gjort nedanstående tabell. Den är uppställd efter Gibbard & von Kolfschoten ingående analys i boken A Geologic Time Scale från Cambridge University Press.
I tabellen är många dateringar ungefärliga. Det beror till en del på att det oftast är oklart när en glaciation eller interstadial exakt började. En inlandsis kunde under flera tusen år byggas upp i ett område utan att geologerna kan lägga märke till det för att sedan spridas över ett vidare område då den blir tydligt observerbar. Ett annat problem är att geologerna använder olika termer i olika områden: En terminologi för Nordamerika, en för Storbritannien, en för Tyskland, en för Schweiz och en för Ryssland m fl. Jag har i tabellen har följt benämningarna för Tyskland eller egentligen Nordvästeuropa.
| Istiderna under senaste 2,6 miljoner åren | ||
| 115 000 år sedan | 130 000 år sedan | 325 000–130 000 år sedan |
|---|---|---|
| Weichsel glaciationen började | Eem interglacialen började | Tiden för Saale glaciationen |
| Cirka 140 000 år sedan | Cirka 150 000 år sedan | Cirka 200 000 år sedan |
| Warthe maximumet | Sista Drenthe maximumet kulminerade följt av den korta Seyda interstadialen |
En serie på 3 stycken Drenthe maxima började |
| Cirka 250 000 år sedan | Cirka 270 000 år sedan | Cirka 325 000 år sedan |
| Wacken glaciationen | Dömnitz interglacialen | Fuhne glaciationen Början på Saale istiden |
| 424 000–374 000 år sedan | 400 000–340 000 år sedan | 866 000–478 000 år sedan |
| Holstein interglacialen | Elster glaciationen | Cromer komplexet med tre maxima och fyra interstadialer |
| 1,140 000–880 000 år sedan | 1,28 milj år sedan | 1,5 milj år sedan |
| Baveliska glaciationen med fyra maxima |
Menap glaciationen | Waaliska glaciationen |
| 1,78 milj år sedan | 2,3 – 1,8 milj år sedan | 2,5 milj år sedan |
| Euboriska glaciationen | Tiglien komplexet med fem 40 000 år långa glaciationer Torka i Östafrika Sahara öken |
Praetiglien interglacialen |
| 2,6 milj år sedan | ||
| Reuvian C glaciationen Antarktis täcks med inlandsis Torka i Östafrika Sahara öken |
||
Borrningar i Adenviken
I Adenviken, vattnen mellan Jemen på Arabiska halvön och Norra Somaliland i Afrika, har gjorts borrningar i havsbottnen. Borrkärnorna har avslöjat att Östafrika växlade mellan torrt och vått klimat i en regelbunden takt av 23 000 år (deMenocal 2014) sedan åtminstone 1,4 miljoner år tillbaka. Enligt den amerikanska maringeologen Peter deMenocal vid Columbiauniversitet i New York, som var med om borrningarna i Adenviken, skulle takten ha samband med jordens precession, d v s att jordaxeln lutar lite extra, en lutning som gör ett varv på en tid av 25 800 år (nordpolen pekar åt olika håll under denna tid). Men tidsspannen är inte helt identiska, varför man kan misstänka att även väderfenomenet El Niño var inblandat. Men de häftiga kasten mellan våta och torra perioder var en del av en process som ledde till att savannen bredde ut sig alltmer på de tidigare skogarnas bekostnad i Östafrika.
deMenocal framhåller att under två perioder var savannens framryckning särskilt framträdande: Den ena inträffade för mellan 2,9-2,6 miljoner år sedan, den andra för mellan 1,9-1,6 miljoner år tillbaka (deMenocal 2011). Vid den första försvann Australopithecus afariensis och i stället dök Paranthropus bosei och P robustus upp. Den berömda Lucy tillhörde A afariensis och levde till stor del i träd och på dess mångahanda blad och frukter. Och när träden försvann, dog hon ut. Till det nya savannlandskapet anpassade sig Paranthropustyperna som en ny, marklevande australopitecin. Paranthroperna hade kraftiga käkar och effektivt malande tänder, som därmed kunde utnyttja savannens svårtuggade vegetabiliska näringsresurser (ibid). Vid den andra uppenbarade sig Homo erectus/ergaster. Även hon var marklevande men hade den dittills största hjärnan av homininerna. Hon kunde eller lärde sig springa och levde i mera komplicerade grupper än australopitecinerna. Framförallt kunde hon vid sidan om växtföda konsumera kött från slagna eller självdöda djur (ibid). Hon var också den första homininen som lämnade Afrika (och kallas H erectus i Eurasien). I Afrika uppfann den kvarvarande H ergaster stensmidet av acheultyp.
Men även under Homo ergasters fortsatta liv på savannen fortsatte svängningarna mellan vått och torrt. Varje plötsligt omslag efter stabilt klimat i 23 000 år gav sannolikt upphov till en klimatchock hos henne. Varje gång chocken kom, alltid efter ett så stort antal generationer att förra chocken glömts bort, hade arvsanlagen svårt att hänga med. Långsamt tvingades de släppa den hårda styrning, som de sannolikt hade i fråga om Paranthroperna (vilket småningom ledde till deras död). Men i fråga om H ergaster (och förmodligen också H erectus utanför Afrika) medförde klimatchockerna att genomet släppte något av sitt grepp. Hur det gick till vet vi inte. Men en indikation är att åldern för vuxenblivandet långsamt försköts. Australopitecinerna blev troligen fullvuxna i tolvårsåldern som schimpanserna blir det än i dag. Den tidiga Homo ergaster blev det också som framgick av beskrivningen av Turkanapojken i kapitel 2. Men hos den sena H ergaster är det mycket möjligt att åldern hade börjat förskjutas till 15 år. Homo heidelbergensis, som började tala ett fonologiskt språk för cirka 1 miljon år sedan, skulle ha blivit fullvuxen vid 15 år. En halv miljon år senare skulle Mauermannen småningom ha fått sin barn- och ungdom förlängd till 18-årsålder. Fram till dess hade det individuella spelrummet varit genetiskt ganska begränsat, kanske stod 10-15 procent av beteendet utanför genernas kontroll. Med Mauermänniskorna började det öka så att hos de moderna människorna 27-28 procent är fritt.
En annan indikation kan sannolikt vara det fonologiska språkets framväxt. Den neurologiska evolution, som krävdes för att åstadkomma de serier av ljud, som kan sammanställas till ord, måste ha krävt en förhållandevis lång tid. Som Genevieve Konopka och hennes team från Texasuniversitets sydvästra medicinska center, där man forskar om autism, visade i en artikel år 2009, är den genetiska komplikationen avsevärd. Inte mindre än 107 gener måste kopplas in innan den mänskliga talapparaten fungerar (Konopka et al 2009). Central är därvid FOXP2-genen i kromosom 7 (se bilden nedan).
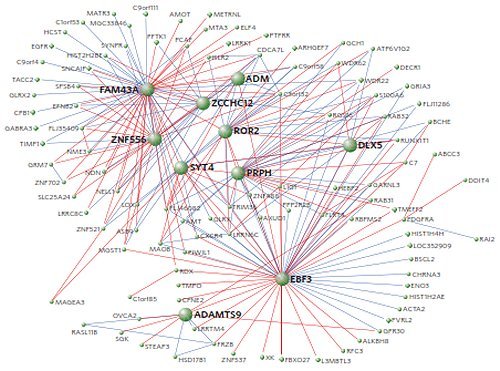
Visualisering av de genetiska moduler i kromosom 7, där språkgenen FOXP2 hos människan och FOXP2CHIMP hos schimpansen finns. På bilden visas 250 gener och hur de är relaterade till andra gener. Samma gener hos människa och schimpans är tecknade i rött, de som endast utmärker människan i blått. De gener som spelar rollen av styrande center har fet stil. Schema efter Konopka et al 2008.
Fram och tillbaka
För att ett så omfattande genetiskt kopplingssystem ska växa fram krävs tid. Det är därför mycket möjligt att det faktum att Homo ergaster verkar ha försvunnit kring för 1,4 miljoner år sedan och de första lämningarna av Homo heidelbergensis dyker upp kring 1 miljon år tillbaka har ett samband. Vi kan misstänka att isolerade grupper av H ergaster i Östafrika vandrade från kusten inåt land under varje 23 000 år långa våtperiod för att sedan tvingas söka sig till kusten när det torra skedet kom tillbaka. Dessa återkommande miljöbyten kan ha drivit fram de omfattande förändringar av gener och nervsystem (se nästa kapitel), som ledde till det fonologiska språket uppkomst. Utvecklingen skulle ha varit fullbordad med H heidelbergensis framträdande.
Som vi såg i förra filen, så inträffade en liknande klimatologisk process i Nordafrika. Men där var Sahara vattenrik och grönskande under de varma och våta perioderna. Under de torra och kalla tiderna förvandlades den åter till öken. Även om inga bevis ännu föreligger där det fullt möjligt att H ergaster kunde spridas ut över Sahara under de förstnämnda perioderna för att drivas ut mot kusterna i nordväst och mot skogarna i söder under de senare. De fynd vi har i dag pekar på att det kunde ha varit i Nordafrika som H ergaster utvecklades till H heidelbergensis. Men kan det ha skett en utveckling parallellt i Nordafrika och Syd/Östafrika?
Det är som sagt i Afrika som H heidelbergensis lämnade sin första fossila spår. Detta förhållande öppnar för nya frågor. Var det enbart hos den sena H ergaster som det fonologiska språket steg för steg växte fram? Om det var fallet var H erectus, som hade hunnit sprida ut sig över Eurasien, visserligen inte stum, men skulle bara ha haft ett enkelt språk sammansatt av olika slag av varningsrop? Kan det ha förhållit sig så att när H heidelbergensis för cirka 700 000 år sedan kom till Europa spred hon sina gener för fonologiskt språk till H antecessor? Hon började då först tala på ett modernt sätt. Eller hade den hittills okända antropologiska typ som H erectus/ergaster växte fram ur, redan anlag för tidiga former av ett enkelt fonologiskt språk? Det är onekligen spännande att vi kan börja ställa sådana frågor!
Hjärnans och kroppens storlek
Som synes är alla detaljer i den process som ledde till att fonologiskt språk uppstod ännu inte helt förstådda. I det följande ska jag ta fram en del infallsvinklar som bidrar att synliggöra sammanhanget. Det finns en aspekt på evolutionen som är förvånansvärt upplysande, nämligen förhållandet mellan hjärnvolym och kroppsstorlek. Av landdjuren har elefanterna de största hjärnorna men också den största kroppsvolymen. Hur är det med de tidiga homininerna i jämförelse med schimpanserna? Tack vare ett samarbete mellan universiteten i Cambridge i England och Tübingen i Tyskland har utarbetats en metod för mätande av längd och vikt, alltså kroppsvolymen, på de tidiga homininerna. Man har t o m kunnat utnyttja så små fossiliserade benbitar som tår. Resultatet visar att australopitecinerna och de tidiga homo var alla småväxta. Det var sedan med H ergaster som en långsam ökning av längd och kroppsvikt skedde (Will & Stock 2015). Detaljerna framgår av tabellen nedan. Tabellen går kronologiskt nerifrån uppåt.
| Fynd- platser |
Medel- längd |
Mini- längd |
Maxi- längd |
Medel- vikt |
Mini- vikt |
Maxi- vikt |
Hjärn- volym |
| Koobi Fora (Kenya) mindre än 1,7 milj år sedan |
166,2 cm | 156,7 cm | 178 cm | 71 kilo | 60 kilo | 81 kilo | Homo ergaster: 700-900 cm3 |
| Sydafrikanska grottor, 1,8-1,4 milj år sedan |
149,1 cm | 135,8 cm | 159,4 cm | 45,5 kg | 32 kg | 62 kg | Australo- pithecus sediba: 420-450 cm3 |
| Olduvai (Tanzania) 1,75 milj år sedan |
146,9 cm | 133,8 cm | 161,7 cm | 50 kg | 38 kg | 65 kg | Australo- pithecus boisei: 500-550 cm3 |
| Dmanisi (Georgien) 1,8 milj år sedan |
149,7 cm | 146,7 cm | 152,7 cm | 49,5 kg | 46 kg | 50 kg | Dmanisi 5: 546 cm3 |
| Koobi Fora (Kenya) mer än 1,8 milj år sedan |
154,7 cm | 147,9 cm | 168,1 cm | 56,4 kg | 39 kg | 69 kg | Homo habilis: 550-687 cm3 |
| Källor: Will & Stock 2015, hjärnvolymerna från Wikipedia på respektive uppslagsord. | |||||||
Tabellen går nerifrån uppåt. De tidiga homininerna var kortare och lättare än schimpanserna, men de hade större hjärnor. Schimpanserna har en genomsnittlig längd på 1,7 m och väger omkring 70 kg. Hjärnvolymen spänner över 282-500 cm3. Det är först med H ergaster som en hominin når en kroppslängd och -vikt jämförbar med schimpansernas. Men då är hjärnvolymen den dubbla hos henne.
Australopitecinerna intelligentare?
Schimpanserna har en genomsnittlig längd på 1,7 m och väger omkring 70 kg. Hjärnvolymen spänner över 282-500 cm3 (källa Wikipedia 2015-03-18). De tidiga homininerna var kortare och lättare än dagens schimpanser. Men de hade större hjärnor. Det är först med H ergaster som en hominin når en kroppslängd och -vikt jämförbar med schimpansernas. Men då är hjärnvolymen den dubbla hos henne. Att döma av dessa skillnader skulle de sena australopitecinerna och de tidiga homo ha varit klart intelligentare – eller i varje fall proportionellt hade större hjärnor – än schimpanserna.
Australopitecinernas större hjärnor kan förklaras med den tvåbenta gången, bipedalismen. Den kräver ett omfattande neurologiskt maskineri för att fungera. Men frågan är vilka andra inslag kan ha stimulerat hjärnutvecklingen.
Ett förslag till förklaring kom i maj 2015 i en artikel i vetenskapstidskriften Nature. Där beskrevs fynd av stenverktyg – vässade skärstenar, hammarstenar och stenar som använts som städen –, som gjorts på västra stranden av Turkanasjön i norra Kenya. Men det anmärkningsvärda med dessa verktyg var deras ålder: 3,3 miljoner år (Harmand et al 2015). Detta smide är 700 000 år äldre än Olduwan-smidet, som anses ha uppfunnits av Homo habilis. Det har döpts till Lomekwian-smide (ibid). Det är ett smide som någon australopitecin-art har utvecklat. Det sägs påminna om schimpansernas sätt att tillverka verktyg, men är lite mera avancerat (ibid) – något som man kunde vänta sig av en större hjärna!
Nära till orangutangerna
Ett annat nytt perspektiv på den moderna människans utveckling kom år 2011 tack vare en stor undersökning av orangutangernas genom, utförd av den medicinska fakulteten vid Washington-universitetet i Washington, USA, under ledning av Devin P Locke. Den visade sensationellt att 97 procent av vårt DNA är identiskt med orangutangernas (Locke et al 2011). Människans och orangutangernas DNA står de facto närmare den tradition av apor, som förlorade sin svans för kanske 20 miljoner år sedan, än gorillor och schimpanser. Deras DNA har utvecklats i samband med den anpassning till de nischer, som de lever i än i dag.
Det märkliga förhållandet att gibboner och människor kan sjunga och orangutanghannar bär skägg som männen gör, två drag som hittills varit oförklarliga och därför nonchalerade, har med undersökningen från Washington-universitetet äntligen fått en förklaring: Människan har bevarat ”ålderdomliga” drag därför att de inte sorterats bort av utvecklingen genom en anpassning till en särskild nisch.
Att vi har så mycket mera orangutang i våra gener än någon tidigare tänkt sig, har än så länge inte riktigt absorberats av paleoantropologerna. Men det visar Russell Wallaces tankar att orangutangerna och människorna står nära varandra inte är helt fel. Människan växte fram i Afrika, som Darwin föreslog, men inte nödvändigtvis med gorillor och schimpanser som mellanstationer. Kanske Björn Kurtén rentav får sina kätterska tankar om att människan inte utvecklades från aporna utan följde en egen linje, framlagd i boken Inte från aporna (Kurtén 1971), också får en revansch, åtminstone delvis.
Det ser nämligen i dagens kunskapsläge inte bättre ut än att människans utveckling började med en svanslös apa för mer än 20 miljoner år sedan. Från den växte fram en apart som levde i Europa och Västasien, Pliobates cataloniae. Den skulle ha varit förfader till både gibboner och dagens högre apor (Alba et al 2015). Den skulle ha förblivit liten och trädlevande, men kom att avsätta, större arter som dels levde i träd, dels på marken. En av de möjliga arvtagarna i Europa var Danuvius guggenmosi, vars fossil nyligen hittades i Bayern (se kapitel 1). Utanför Europa har gjorts flera fynd, som kan var relaterade till Pliobates. En av dem var Ankarapithecus, som levde för 18 miljoner år tillbaka i Mindre Asien (Alpapgut et al 1996). I södra Asien växte åter fram en annan marklevande typ, nämligen Sivapithecus för 14 miljoner år sedan. Den kom småningom att bli orangutang (Pilbeam 1982). Men den lilla och trädlevande versionen skulle ha vandrat till Afrika över en landbro från Arabiska halvön till Etiopien (Röda havet var då ännu en vik av Medelhavet). Där utvecklades hon till de tidiga, trädlevande australopiteciner, som emellertid gick på marken mellan träden på två ben (som gibboner och delvis orangutanger). Gorillor och schimpanser är marklevande och går på fyra ben. De stora avvikelser från denna huvudlinje som förde till människa för 10 respektive 7 miljoner år tillbaka. Vi får emellertid se hur bilden kommer att framträda när Washington-universitetets forskningsresultat diskuterats färdigt bland fackmännen.
Evolutionens ”experiment”
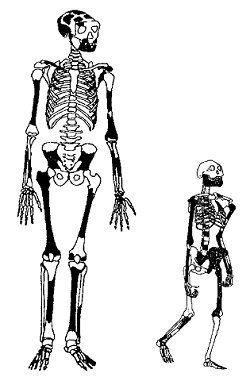
Rekonstruerade skelett i skalenlig storlek av två kvinnor från människans utvecklingsträd med ett avstånd i tid mer än en miljon år. Till höger har vi det rekonstruerade skelettet av Australopithecus afariensis, också känd som Lucy, cirka 3,2 miljoner år gammalt. Lägg märke till de långa armarna. Till vänster det rekonstruerade skelettet från fyndet ER 1808 från Koobi Fora, Kenya, 1,7 miljoner år gammalt. När Homo erectus trädde fram för cirka 2 miljoner år sedan var hon ungefär lika liten som Lucy. Men sedan växte hon snabbt i storlek i en anpassning till ett liv på slätterna. Notera höfterna och de långa benen, som visar att hon hade lärt sig löpa. Med svart är markerat de bevarade fossilfynden. Bild efter Hawks et al 2000.
Men vad föranledde Homo ergasters hjärntillväxt? Var det klimatchocker, som tvingade fram en ökad förmåga till anpassning, bl a att kunna tillgodogöra sig vad havet kunde erbjuda i matväg?
Det kan ha samband med att utvecklingen tycks ha drivits fram som ett alternativ av evolutionen själv genom ”experiment”. Detta föreslogs i en tolkning av evolutionen, som presenterades i vetenskapstidskriften Science år 2014 av Susan C Antón från New York-universitetet i New York, USA. När klimatchockerna gjorde miljön oförutsägbar, tog evolutionen fram olika lösningar, alltså begick ”experiment” (Antón et al 2014). Den typ av australopitecin som kunde tillgodogöra sig så många matkällor som möjligt, som alltså var bredast i sin diet, kom att bli Homo ergaster (ibid). I sitt svar på klimatchockerna skulle redan australopitecinerna ha börjat en utveckling mot optimal adaptabilitet (alltså bästa möjliga förmåga till anpassning).
Nu har t o m preciserats ett förslag var evolutionens ”verkstad” skulle ha legat, nämligen Turkana-bassängen i Kenya. Professorn i evolutionär paleontologi vid Helsingfors universitet, Mikael Fortelius, har med ett team undersökt alla fynd av fossiliserade tänder från växtätande djur som gjorts i bassängområdet. Han har kunnat följa kasten mellan vått och torrt klimat ända upp till för 8 miljoner år sedan. Han presenterade resultaten från undersökningen i ett föredrag, som han höll vid ett möte i oktober 2015 vid Royal Society i London, en sammankomst till ära för paleoantropologen Richard Leakey på dennes 70-årsdag. Enligt Fortelius skulle området kring Turkana-bassängen genom tiderna ha tenderat reagera först vid klimatomslag. De djur som anpassade sig till det nya torra klimatet kunde sedan spridas över större områden när torkan bredde ut sig över vidare områden. Bassängen skulle ha varit en ”fabrik för nya arter” (Wong 2015).
Men om de sena H ergaster levde i bassängen, stannade de inte kvar när torkan började kännas av. De bröt upp och flyttade till kusten. Där behövde de varje gång de nya ord för att beskriva sin nya miljö. Det kan ha drivit på uppkomsten av det fonologiska språket.
I nästa kapitel ska vi diskutera hur detta språk kan ha sett ut hos H heidelbergensis.
Läs även essäns sjunde kapitel om Heidelbergensarens språkförmåga.
Till historiemenyn.
Källor till detta kapitel:
Alba et al 2015: David M. Alba1, Sergio Almécija, Daniel DeMiguel, Josep Fortuny, Miriam Pérez de los Ríos, Marta Pina, Josep M. Robles, Salvador Moyà-Solà: Miocene small-bodied ape from Eurasia sheds light on hominoid evolution. Science 30 October 2015, vol. 350, no. 6260.
Alpagut et al 1996: Berna Alpagut, Peter Andrews, Mikael Fortelius, John Kappelman, Ilhan Temizsoy, Hürkan Çelebi & William Lindsay: A new specimen of Ankarapithecus meteai from the Sinap Formation of central Anatolia. Nature 382 (25 July 1996).
Antón et al 2014: Susan C. Antón, Richard Potts, Leslie C. Aiello: Evolution of early Homo: An integrated biological perspective. Science 4 July 2014, vol. 345, no. 6192.
Darwin 1859: Charles Darwin: Om arternas uppkomst. Svensk översättning av Roland Adlerbert och Lars-Ove Skeppsholm. Bonniers 1980. Orginalets titel On the Origin of Species by Means of Natural Selection, utgiven 1859.
deMenocal 2011: Peter B. deMenocal: Climate and Human Evolution. Science, 4 february 2011, vol 331.
deMenocal 2014: Peter B. deMenocal: Klimatshocks. Scientific American, September 2014.
Gibbard & von Kolfschoten 2004: P. Gibbard and T. van Kolfschoten: The Pleistocene and Holocene Epochs. Chapter 22 in A Geologic Time Scale 2004 Edited by Felix M. Gradstein, James G. Ogg, and Alan G. Smith. Cambridge university Press.
Harmand et al 2015: Sonia Harmand, Jason E. Lewis, Craig S. Feibel, Christopher J. Lepre, Sandrine Prat, Arnaud Lenoble, Xavier Boës, Rhonda L. Quinn, Michel Brenet, Adrian Arroyo, Nicholas Taylor, Sophie Clément, Guillaume Daver, Jean-Philip Brugal, Louise Leakey, Richard A. Mortlock, James D. Wright, Sammy Lokorodi, Christopher Kirwa, Dennis V. Kent & Hélène Roche: 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature 521, 310-315 (21 May 2015).
Hawks et al 2000: John Hawks, Keith Hunley, Sang-Hee Lee, and Milford Wolpoff: Population Bottlenecks and Pleistocene Human Evolution. Mol. Biol. Evol. 17(1):2-22. 2000.
Konopka et al 2009: Genevieve Konopka, Jamee M. Bomar, Kellen Winden, Giovanni Coppola, Zophonias O. Jonsson, Fuying Gao, Sophia Peng, Todd M. Preuss, James A. Wohlschlegel, and Daniel H. Geschwind: Human-specific transcriptional regulation of CNS development genes by FOXP2. Nature 462, 12 November 2009.
Kurtén 1971: Björn Kurtén: Inte från aporna.
Lai et al 2016; Shenshen Lai, Javad Safaei, Steven Pelech: Evolutionary ancestry of eukaryotic protein kinases and choline kinases. Journal of Biological Chemistry, 2016; jbc.M115.691428 DOI: 10.1074/jbc.M115.691428
Locke et al 2011: Devin P. Locke och inte mindre än 99 medarbetare: Comparative and demographic analysis of orangutan genomes. Nature Vol. 469, 27 January 2011.
Pilbeam 1982: David Pilbeam: New hominoid skull material from the Miocene of Pakistan. Nature 295 (21 January 1982).
Ravila 1967: Paavo Ravila: Totuus ja metodi - Kielitieteellisiä esseitä. (Sanningen och metoden - språkvetenskapliga essäer). WSOY.
Will & Stock 2015: Manuel Will and T. Stock: Spatial and temporal variation of body size among early Homo. Journal of Human Evolution, 2015 DOI: 10.1016/j.jhevol.2015.02.009
Wong 2015: Sam Wong: Did climate change jump-start human evolution in East Africa? New Scientist, 4 November 2015.
Publiceringshistoria: Utlagd 15-10-30. Uppdaterad 16-04-04. Omarbetad och ånyo uppdaterad 18-12-20. Ytterligare omarbetad och layouten förnyad 20-08-10.
Sänd en kommentar till essän!
Bilder och texter får inte lånas utan tillstånd. Citat ur texter är tillåtet med angivande av källan.